THZ1
更新时间:2025-08-24 19:55:13
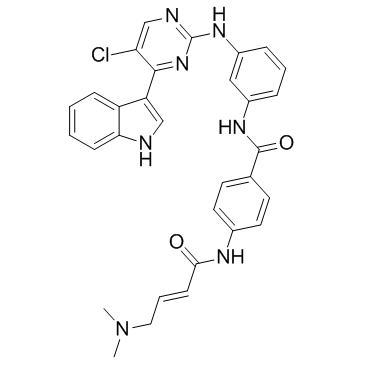
THZ1结构式

|
常用名 | THZ1 | 英文名 | THZ1 2HCl |
|---|---|---|---|---|
| CAS号 | 1604810-83-4 | 分子量 | 566.053 | |
| 密度 | 1.4±0.1 g/cm3 | 沸点 | N/A | |
| 分子式 | C31H28ClN7O2 | 熔点 | N/A | |
| MSDS | N/A | 闪点 | N/A |
THZ1用途THZ1 是有效,选择性的共价 CDK7 抑制剂,IC50 为 3.2 nM。 |

| 中文名 | N-[3-[[5-氯-4-(1H-吲哚-3-基)-2-嘧啶基]氨基]苯基]-4-[[(2E)-4-(二甲基氨基)-1-氧代-2-丁烯-1-基]氨基]苯甲酰胺 |
|---|---|
| 英文名 | N-[3-[[5-Chloro-4-(1H-indol-3-yl)-2-pyrimidinyl]amino]phenyl]-4-[[(2E)-4-(dimethylamino)-1-oxo-2-buten-1-yl]amino]benzamide |
| 英文别名 | 更多 |
| 描述 | THZ1 是有效,选择性的共价 CDK7 抑制剂,IC50 为 3.2 nM。 |
|---|---|
| 相关类别 | |
| 靶点实验 |
CDK7:3.2 nM (IC50) |
| 体外研究 | THZ1抑制Jurkat细胞和Loucy细胞,IC50分别为50 nM和0.55 nM。 THZ1显示体外CDK7的时间依赖性抑制和细胞内CDK7的共价结合。 THZ1(9,27,83,250,750和2500nM)抑制CDK12,但与CDK7相比具有更高的浓度。 THZ1(1μM)不可逆地抑制RNAPII CTD和CAK磷酸化。 THZ1(2.5μM)通过共价靶向位于Hela S3细胞中CDK7激酶结构域外的独特半胱氨酸,不可逆地抑制RNAPII CTD磷酸化。 THZ1(250 nM)导致细胞增殖减少和细胞凋亡指数增加,同时抗凋亡蛋白减少,最显着的是T-ALL细胞系中的MCL-1和XIAP [1]。低剂量THZ1(50 nM)治疗引起食管鳞状细胞癌(OSCC)中许多致癌转录物的选择性抑制[2]。所有基因型不同的人(hSCLC)细胞系对THZ1表现出高度敏感性,IC50在5-20nM范围内[3]。 |
| 体内研究 | THZ1(10 mg/kg)显示有效杀死原发性慢性淋巴细胞白血病(CLL)细胞和抗原发性TALL细胞的抗增殖活性,并在体内对抗人T-ALL异种移植物[1] .THZ1(10 mg/kg,ip )完全抑制体内食管鳞状细胞癌肿瘤生长而不丧失体重或其他常见毒性作用[2]。 THZ1(10 mg/kg,iv)抑制人MYCN扩增NB小鼠模型中的肿瘤生长,并且没有毒性[4]。 |
| 激酶实验 | 对于来自293A细胞裂解物的HCT116或FLAG-CDK12的FLAG-CDK7蛋白免疫沉淀后的激酶测定,首先用THZ1,THZ1-R或DMSO在37℃处理细胞4小时。然后通过在50mM Tris HCl pH 8.0,150mM NaCl,1%NP-40,5mM EDTA和蛋白酶/磷酸酶混合物中裂解来收获细胞。使用FLAG抗体 - 缀合的琼脂糖珠从细胞裂解物中免疫沉淀外源CDK7或CDK12蛋白。沉淀的蛋白质用裂解缓冲液洗涤6次,然后用激酶缓冲液(40mM Hepes pH 7.5,150mM NaCl,10mM MgCl 2,5%甘油)洗涤2次,并在30℃下进行体外激酶测定45分钟。使用1μg大的RNAPII亚基(RPB1)作为底物和25μMATP和10μCi的32P ATP。使用重组CDK7 / TFIIH / MAT1的激酶测定以上述方式使用每次反应25ng CAK复合物进行。对于设计用于测试CDK7激酶活性的时间依赖性失活的激酶测定,将CAK复合物与指定浓度的THZ1,THZ1-R或DMSO在不含ATP的激酶缓冲液中在30℃下预孵育4小时,然后进行激酶测定条件[1]。 |
| 细胞实验 | 将Jurkat,Loucy,KOPTK1和DND-41细胞系接种于384孔微量培养板中,在含有5%FBS和青霉素/链霉抗生物素蛋白的培养基中以15%汇合。用THZ1(2,10,50,250,1250和6250nM)或DMSO处理细胞72小时,并使用刃天青[1]测定细胞活力。 |
| 动物实验 | 小鼠[1] 32只NOD-SCIDIL2Rcγnull(NSG)9周龄雌性小鼠根据平均BLI分为治疗组,如下:THZ1 10 mg / kg qD,THZ1 10 mg / kg BID和载体(10%) D5W中的DMSO)BID(所有组n = 10)。排除两只小鼠,一只具有最高,一只具有最低的BLI。所有治疗均通过侧尾静脉中的IV注射以3.3μL/ g(非盲)的体积施用。对小鼠进行成像并每3-5天称重一次。处理小鼠四周,并在最后一天对小鼠进行成像,给药并在给药后约5-6小时处死。处死后,在EDTA管中通过心脏穿刺收集血液;处理一部分(300μL)用于等离子体。从每只小鼠收集肝脏和脾脏组织,每个样品的一半快速冷冻,每个样品的一半固定。处理血浆和肝脏样品用于THZ1的药代动力学分析。将脾组织匀浆并裂解并处理以用于THZ1靶接合的药效学分析。 |
| 参考文献 |
| 密度 | 1.4±0.1 g/cm3 |
|---|---|
| 分子式 | C31H28ClN7O2 |
| 分子量 | 566.053 |
| 精确质量 | 565.199280 |
| LogP | 5.08 |
| InChIKey | OBJNFLYHUXWUPF-IZZDOVSWSA-N |
| SMILES | CN(C)CC=CC(=O)Nc1ccc(C(=O)Nc2cccc(Nc3ncc(Cl)c(-c4c[nH]c5ccccc45)n3)c2)cc1 |
| 外观性状 | 粉末 |
| 折射率 | 1.735 |
| 储存条件 | -20℃ |
| N-(3-{[5-Chloro-4-(1H-indol-3-yl)-2-pyrimidinyl]amino}phenyl)-4-{[(2E)-4-(dimethylamino)-2-butenoyl]amino}benzamide |
| THZ1 |
本网页内容来自不同专业数据源,如对内容有疑义,欢迎联系service1@chemsrc.com。

我想出现在这里:
 351666998
复制
351666998
复制

 13311869306
复制
13311869306
复制

- 上海阿拉丁生化科技股份有限公司
- 上海市 上海市 浦东新区
- 产品名:THZ1 2HCl
- 纯度:98.0%
- 价格: ¥5973.9/100mg ¥926.9/10mg ¥2059.9/25mg ¥3295.9/50mg
- 联系人:阿拉丁
 2881459338
复制
2881459338
复制
- 联系电话:400-620-6333
- 广东翁江化学试剂有限公司
- 广东省 韶关市 翁源县
- 产品名:THZ1
- 纯度:98.0%
- 价格: ¥1000.0/5mg
- 联系人:朱丽丹
13927877242
复制
 13927877242
复制
13927877242
复制

- 联系电话:13927877242
- 上海化源世纪贸易有限公司
- 上海市 上海市 普陀区
- 产品名:THZ1
- 纯度:98.0%
- 价格:
- 联系人:徐经理
351666998
复制
13311869306
复制
- 联系电话:13311869306
- 武汉壹加壹生物科技有限公司
- 湖北省 武汉市 江夏区
- 产品名:N-[3-[[5-氯-4-(1H-吲哚-3-基)-2-嘧啶基]氨基]苯基]-4-[[(2E)-4-(二甲基氨基)-1-氧代-2-丁烯-1-基]氨基]苯甲酰胺
- 纯度:98.0%
- 价格: ¥需询单/1g ¥需询单/5g
- 联系人:刘经理
673003916
复制
16602739303
复制

- 联系电话:16602739303
查看所有供应商和价格请点击: