IWP 2
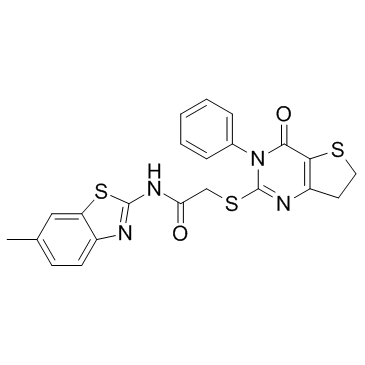
IWP 2结构式

|
常用名 | IWP 2 | 英文名 | IWP-2 |
|---|---|---|---|---|
| CAS号 | 686770-61-6 | 分子量 | 466.599 | |
| 密度 | 1.5±0.1 g/cm3 | 沸点 | N/A | |
| 分子式 | C22H18N4O2S3 | 熔点 | 257 °C(dec.) | |
| MSDS | 美版 | 闪点 | N/A |
IWP 2用途IWP-2 抑制 Wnt 加工和分泌,IC50 为 27 nM。 |

| 中文名 | N-(6-甲基-2-苯并噻唑基)-2-[(3,4,6,7-四氢-4-氧代-3-苯基噻吩并[3,2d]嘧啶-2-基)硫基]乙酰胺 |
|---|---|
| 英文名 | N-(6-Methyl-2-benzothiazolyl)-2-[(3,4,6,7-tetrahydro-4-oxo-3-phenylthieno[3,2-d]pyrimidin-2-yl)thio]-acetamide |
| 英文别名 | 更多 |
| 描述 | IWP-2 抑制 Wnt 加工和分泌,IC50 为 27 nM。 |
|---|---|
| 相关类别 | |
| 靶点实验 |
IC50: 27 nM (Wnt)[1] |
| 体外研究 | IWP-2,WNT加工和分泌的抑制剂。 IWP-2显着增强LEF的抗增殖作用。同样明显的是,与单一药物相比,LEF和IWP-2的组合可以最大程度地降低β-连环蛋白,c-Myc,细胞周期蛋白D1,Bcl2和Bax的表达[2]。在MKN28细胞系中处理4天后,10-50μMIWP-2显着抑制MKN28细胞的增殖(P <0.05)。此外,IWP-2治疗后锚定依赖性和锚定独立的集落数显着降低(P <0.05)[3]。 |
| 体内研究 | 为了评估IWP-2在体内的功效,在注射相似体积的蓝色染料填充的乳胶珠之前约2小时,将200μL的IWP-2-脂质体或游离脂质体分别腹膜内注射到C57BL/6小鼠中。或大肠杆菌DH5α。 IWP-2引起蓝珠和大肠杆菌摄取的显着降低,如通过腹膜灌洗细胞中的CFU在2小时内评估的。此外,与对照值相比,相应小鼠的灌洗液中TNF-α和IL-6的水平降低了2-4倍。有趣的是,IWP-2甚至诱导抗炎细胞因子IL-10的分泌显着增加[4]。用IWP-2预处理显着(P <0.05)消除了SP诱导的Wnt3a,p-GSK3β和β-连环蛋白表达的增加[5]。 |
| 细胞实验 | 将人RCC细胞系786O和Caki-2(5×103)接种到96孔板中。在将Caki-2乙醇与不断增加浓度的LEF和20μMIWP-2一起孵育48小时后,通过MST测定估计细胞活力。处理后,将10μLMTS加入每个孔中孵育2小时。使用型号ELX800 Micro Plate Reader在490nm处测量吸光度。对于集落形成测定,将Caki-2细胞用胰蛋白酶消化成单细胞悬浮液,并以1000个细胞/孔接种到新鲜的6孔板中。然后将细胞与所示浓度的LEF一起温育7天。将菌落用无水甲醇固定15分钟,然后用0.1%结晶紫染色20分钟。用PBS洗涤三次后,用数码相机显示直径超过2mm的菌落[2]。 |
| 动物实验 | 小鼠[4]将约3个月大的C57BL / 6小鼠在笼中于23℃,12小时光照/黑暗循环中饲养4至5只。首先用200μL脂质体-IWP2(L1)或脂质体(L)腹膜内(ip)注射小鼠,然后在200μL无菌PBS中用1×108或2×108CFU大肠杆菌注射2小时。 2小时或24小时后,杀死小鼠,并用5mL无菌冰冷的PBS洗涤腹膜腔。将腹膜灌洗液以300×g离心5分钟,将细胞沉淀重悬于RPMI 1640完全培养基中,并将上清液用于细胞因子测定。对于离体实验,如上从正常小鼠分离腹膜吞噬细胞,并且在进行进一步实验之前,将相同数量的细胞在37℃,5%CO 2中培养在培养基中过夜。大鼠[5]使用体重220-280g的成年,雄性和健康Wistar大鼠。将大鼠随机分成如下6组(n = 72,每组12只):( 1)假手术组(S组),(2)I / R组(I / R组),(3)I / R + DMSO组(DMSO组),(4)I / R + IWP组(组IWP),(5)SP组(组SP)和(6)SP + Wnt抑制剂IWP-2组(组SP + IWP)。在组S中连续灌注心脏120分钟。平衡10分钟后,将分离的心脏连续灌注20分钟,然后进行缺血30分钟,然后在组I / R中再灌注60分钟;组DMSO,IWP,SP和SP + IWP用含有0.5mL / L DMSO,10μMIWP(SIGMA-ALDRICH,USA),2.4vol%七氟醚,2.4vol%七氟醚+10μMIWP的KH溶液接受15分钟灌注,分别在I / R之前进行5分钟冲洗。 |
| 参考文献 |
| 密度 | 1.5±0.1 g/cm3 |
|---|---|
| 熔点 | 257 °C(dec.) |
| 分子式 | C22H18N4O2S3 |
| 分子量 | 466.599 |
| 精确质量 | 466.059174 |
| PSA | 156.11000 |
| LogP | 5.25 |
| InChIKey | WRKPZSMRWPJJDH-UHFFFAOYSA-N |
| SMILES | Cc1ccc2nc(NC(=O)CSc3nc4c(c(=O)n3-c3ccccc3)SCC4)sc2c1 |
| 外观性状 | 固体;White to Yellow to Orange powder to crystal |
| 折射率 | 1.787 |
| 储存条件 | Store at +4°C |
| 水溶解性 | DMSO: >5mg/mL |
| 危险品运输编码 | NONH for all modes of transport |
|---|
|
Ligand-independent canonical Wnt activity in canine mammary tumor cell lines associated with aberrant LEF1 expression.
PLoS ONE 9(6) , e98698, (2014) Pet dogs very frequently develop spontaneous mammary tumors and have been suggested as a good model organism for breast cancer research. In order to obtain an insight into underlying signaling mechani... |
|
|
Notch inhibition induces mitotically generated hair cells in mammalian cochleae via activating the Wnt pathway.
Proc. Natl. Acad. Sci. U. S. A. 112(1) , 166-71, (2015) The activation of cochlear progenitor cells is a promising approach for hair cell (HC) regeneration and hearing recovery. The mechanisms underlying the initiation of proliferation of postnatal cochlea... |
|
|
A switch from canonical to noncanonical Wnt signaling mediates early differentiation of human neural stem cells.
Stem Cells 32(12) , 3196-208, (2014) Wnt/β-catenin signaling is essential for neurogenesis but less is known about β-catenin-independent Wnt signals. We show here that Wnt/activator protein-1 (AP-1) signaling drives differentiation of hu... |
| N-(6-Methyl-1,3-benzothiazol-2-yl)-2-[(4-oxo-3-phenyl-3,4,6,7-tetrahydrothieno[3,2-d]pyrimidin-2-yl)sulfanyl]acetamide |
| IWP-2 |
本网页内容来自不同专业数据源,如对内容有疑义,欢迎联系service1@chemsrc.com。

 351666998
复制
351666998
复制

 133 1186 9306
复制
133 1186 9306
复制

- 上海阿拉丁生化科技股份有限公司
- 上海市 上海市 浦东新区
- 产品名:IWP-2
- 纯度:98.0%
- 价格: ¥4119.9/100mg ¥648.9/10mg ¥1462.9/25mg ¥2338.9/50mg
- 联系人:阿拉丁
 2881459338
复制
2881459338
复制
- 联系电话:400-620-6333
- 武汉敬康恩生物医药科技有限公司
- 湖北省 武汉市 东西湖区
- 产品名:IWP-2
- 纯度:98.0%
- 价格: ¥需询单/1g
- 联系人:周经理
1092734927
复制
 15871494362
复制
15871494362
复制

- 联系电话:15871494362
- 上海化源世纪贸易有限公司
- 上海市 上海市 普陀区
- 产品名:N-(6-甲基苯并[d]噻唑-2-基)-2-[(4-氧代-3-苯基-3,4,6,7-四氢噻吩并[3,2-d]嘧啶-2-基)硫基]乙酰胺
- 纯度:98.0%
- 价格:
- 联系人:徐经理
351666998
复制
133 1186 9306
复制
- 联系电话:133 1186 9306
查看所有供应商和价格请点击: